


200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过





激酶锚定蛋白(A-kinase anchor proteins,AKAP)是一个支架蛋白家族,主要作用是将蛋白激酶、磷酸酶、信号终止分子与其相应的底物隔离,从而调节细胞内信号转导[1]。AKAP12属于AKAPs家族,最初是从一名重症肌无力患者中分离出来,曾命名为Gravin,而在啮齿动物中则被称为Src抑制的C激酶底物[2];通过C端两亲性螺旋结合cAMP依赖的蛋白激酶的调节亚单位、蛋白激酶C(protein kinase C,PKC)、蛋白磷酸酶2B、钙调蛋白来调控细胞周期和细胞骨架的结构。此外,AKAP12通过将细胞内信号蛋白组装成特定的复合体来提高时空信号的特异性和效率。

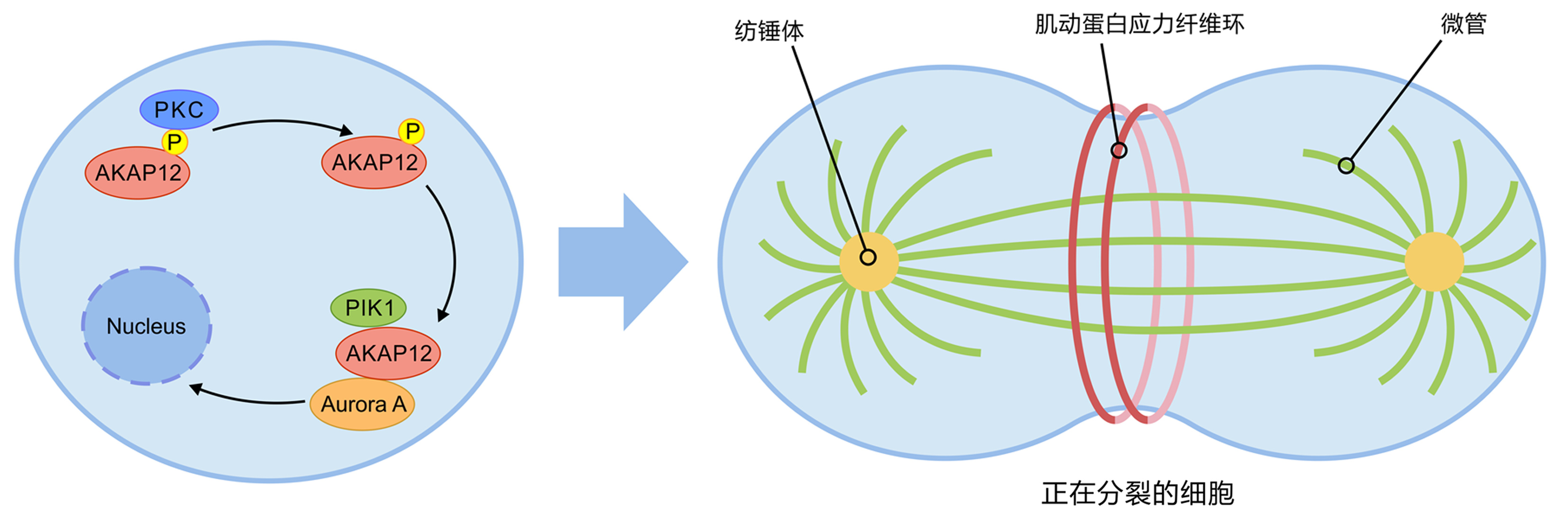
AKAP12是一种有丝分裂负性调节因子,定位于整个细胞质,主要在足小体和细胞边缘富集,在细胞骨架结构调控中起重要作用;经过佛波酯短期处理,AKAP12可从质膜转移到细胞核外围,与此同时,肌动蛋白应力纤维丢失;AKAP12主要引起肌动蛋白的初始溶解,随后应力纤维可重新出现,因此并不抑制肌动蛋白结构的形成。此外,AKAP12与有丝分裂激酶Aurora A和Polo样激酶1形成复合体,在细胞分裂时调节有丝分裂纺锤体的定向,使分裂保持对称性[3]。PKC通过调节AKAP12的丝氨酸磷酸化,改变它与细胞骨架的相互作用以及控制有丝分裂的能力;AKAP12过度表达可诱导细胞扁平化,导致未转化细胞在G1早期发生生长停滞,AKAP12重建肌动蛋白细胞骨架,为后续的细胞分裂做好准备[4-5](图 1)。
图 1 AKAP12调控有丝分裂的过程
Figure 1. The process of AKAP12 regulating mitosis
在血管发育过程中,AKAP12通过调节肌动蛋白细胞骨架调节因子和连接细胞间黏附分子以及肌动蛋白骨架的连接物的表达,在维持血管内皮细胞完整性中发挥重要作用。缺乏AKAP12的斑马鱼胚胎将出现严重出血,部分原因可能是内皮细胞-细胞间粘连紊乱[6]。
此外,AKAP12是调节神经胶质细胞功能的关键因子。在小鼠中,AKAP12在胼胝体中的表达水平高于皮质,并随着年龄的增长而逐渐降低。值得注意的是,AKAP12主要由出生后的星形胶质细胞表达,而不是由胼胝体发育过程中的另一种主要胶质细胞——少突胶质前体细胞表达;与野生型小鼠相比,AKAP12基因敲除小鼠的星形胶质细胞和少突胶质前体细胞数量均减少,出生后胼胝体发育也随之变得缓慢[7]。AKAP12可通过影响血管生成和紧密连接来调节血-脑屏障的形成,通过减少激活蛋白-1,增加星形胶质细胞中抗渗透因子血管生成素1的表达来降低血管内皮细胞生长因子(vascular endothelial growth factor,VEGF)的表达[8];AKAP12还通过增加星形胶质细胞中的血管生成素1含量,降低VEGF水平,促进血-视网膜屏障的形成[9]。
肝纤维化可由多种病因引起,如肝炎病毒感染、酗酒、自身免疫性或代谢性疾病,其特征是胶原和其他细胞外基质过度沉积、瘢痕形成,导致肝脏结构和功能改变[10];近年来,AKAP12在肝纤维化进展过程中的作用越来越受到关注,部分文献报道AKAP12具有抗肝纤维化作用,但也有文献报道,AKAP12发挥促肝纤维化的作用。
经硫代乙酰胺处理8周后,毛细血管化肝窦内皮细胞(liver sinusoidal endothelial cell,LSEC)中AKAP12的表达却呈现下降趋势。AKAP12可减少Ⅰ型胶原α1、弹性蛋白和编码上皮细胞黏附分子的表达,但对细胞外基质的积聚并没有明显影响。随着肝纤维化的进展,AKAP12在门静脉成纤维细胞和LSEC中的表达逐渐减少。在3, 5-二乙氧羰基-1, 4-二氢吡啶(DDC)诱导的胆管纤维化模型中,AKAP12可降低内皮素-1的表达,抑制细胞外基质积聚、血管生成和重塑,阻止门静脉成纤维细胞的激活,促进纤维化消退,并使肌成纤维细胞和肝窦逐渐恢复正常[11]。也有研究[12]表明,AKAP12可以通过抑制PI3K/AKT/PCSK6通路来实现对肝纤维化的抑制作用。
此外,AKAP12也可以通过维护血管内皮屏障功能,保持血管完整性,发挥抗肝纤维化作用。AKAP12可以改善脂多糖诱导的血管内皮细胞功能障碍,降低活性氧和促炎因子的水平,增加内皮型一氧化氮合酶,通过抑制PKC促进细胞外信号调节激酶5的转录活性[13];AKAP12促进内皮损伤修复的机制可能是将PKA和PKC靶向于特定的膜相关部位;以往的研究[14-15]结果表明,PKC有促肝纤维化的作用,PKC抑制剂可减轻肝纤维化,而PKC激活剂——氧化二酰甘油则明显加重CCl4诱导的小鼠肝纤维化;AKAP12在VEGF刺激下,迁移活跃的内皮细胞中表达水平较高,在静止的内皮细胞中表达水平则较低[16]。
组织缺氧是导致肝纤维化进展的另一个重要因素,可诱导AKAP12的表达;AKAP12中有两个低氧诱导因子(hypoxia-inducible factor,HIF)的结合位点,其中最远端的HIF结合位点是缺氧诱导AKAP12的关键。AKAP12以HIF-1α依赖的方式抑制血管生成,增强内皮细胞连接的通透性,可作为血管生成的“刹车”系统。此外,AKAP12还调控PKA激动剂诱导的内皮屏障功能应答[17]。
AKAP12有助于激活肝星状细胞(HSC),HSC是各种肝损伤发生后肝纤维化形成的中心环节。无论是CCl4诱导的急性肝衰竭的肝组织,还是激活的HSC中,AKAP12表达均明显升高,主要影响HSC肌动蛋白骨架的重塑过程[18]。血小板衍生生长因子是一种促进HSC增殖的生长因子,在肝纤维化的进展中起重要作用[19-20],它能够诱导大鼠HSC中的AKAP12酪氨酸磷酸化和细胞骨架网络的快速重排,这是由PKC通过Rho蛋白介导的[21]。同样,在脂多糖诱导的肝损伤中,LSEC中AKAP12的表达显著增加[22]。
上述矛盾的研究结果反映了AKAP12在肝纤维化发展过程中调控的复杂性。
AKAP12在非酒精性脂肪性肝病中的作用也是矛盾的。研究[23]表明,AKAP12在非酒精性脂肪性肝炎(NASH)大鼠模型中表达水平降低,与炎症程度呈负相关,有可能成为其潜在的调控靶点。
但据最近的研究[24]报道,在高脂饲料诱导的动脉粥样硬化小鼠模型中,AKAP12介导的信号复合体通过抑制固醇调节元件结合蛋白-2和肝脏3-羟基-3-甲基-戊二酰辅酶A还原酶的表达,增加主动脉斑块的形成,加重高脂饲料喂养小鼠肝脏中的脂肪堆积和损伤,促进了高脂血症和动脉粥样硬化。
上述研究结果也得出了相反的结论,再次证实了AKAP12调控作用的复杂性,值得进一步研究证实。
AKAP12的磷酸化水平在酒精性肝病/酒精性肝炎患者中普遍升高,通过减少热休克蛋白47(heat shock protein 47,HSP47)与胶原之间的相互作用,AKAP12发挥抑制肝纤维化的作用;乙醇可增加AKAP12的磷酸化水平,降低HSC中AKAP12对PKC/Cyclin-D1的支架活性,诱导HSP47与胶原的相互作用,从而促进HSC活化[25]。
AKAP12是一种肿瘤抑制因子,通过抑制PKC的激活,阻断细胞周期,抑制肿瘤细胞的血管生成、增殖、趋化和侵袭[26],AKAP12在多种肿瘤中的表达下降, 这可能是AKAP12通过其支架功能抑制PKC的激活导致的[27], 启动子超甲基化是多种癌症中AKAP12表达缺失的基础,也是癌症进展和转移的潜在生物标志物。近年来,AKAP12与HCC的关系日益受到关注。AKAP12可能是HCC的候选抑癌基因,在HCC组织中通过启动子高甲基化而下调表达,85%的HCC组织中观察到AKAP12基因启动子的高甲基化,AKAP12 DNA甲基化与HCC患者的低复发风险和长期无进展生存密切相关[28],AKAP12在细胞中的过度表达或缺乏会改变对恶性肿瘤进展或转移的易感性,AKAP12缺陷小鼠的肿瘤发病率更高。
一种新的人类长链非编码RNA(LncRNA)——LNC-HC,在HCC患者中的水平显著降低,LNC-HC可作为竞争的内源性RNA,竞争性结合hsa-miR-183-5p,从而抑制肝癌细胞的增殖,而AKAP12是hsa-miR-183-5p的一种靶基因[29]。miR-103通过抑制AKAP12的表达,增加PKCα的活性,促进肝癌细胞增殖,抑制其凋亡;而AKAP12的过度表达和PKC表达的下调都可以通过抑制端粒酶的活性来抑制肿瘤的生长和转移[30]。HCC组织中AKAP12基因表达下调,并与miR-1251-5p水平呈负相关,AKAP12的过表达降低了肝癌细胞的增殖、迁移和侵袭,而抑制AKAP12则增强了miR-1251-5p基因敲除后减弱的肝癌细胞的增殖、迁移和侵袭,miR-1251-5p通过靶向AKAP12参与抗肿瘤作用[28]。此外,有研究[31]表明,黄连素对人肝癌细胞有明显的细胞毒作用,黄连素在HepG2细胞中的抗增殖作用涉及到细胞凋亡和细胞周期停滞,黄连素调节AKAP12信号通路可能为HCC的治疗提供一个有前景的策略。
AKAP12在慢性肝病中的作用较为复杂,在硫代乙酰胺和DDC诱导的肝纤维化中,毛细血管LSEC中AKAP12的表达减少; 与上述结果矛盾的是,CCl4诱导活化的HSC和脂多糖诱导的LSEC中AKAP12的表达增加;因此,有必要进一步研究AKAP12在肝纤维化发生发展中的作用。AKAP12也参与了NASH的发生和发展,可能是治疗NASH相关炎症的潜在靶点;乙醇通过增加AKAP12的磷酸化和上调HSP47与胶原之间的相互作用来促进酒精性肝病中HSC的激活。作为一种肿瘤抑制因子,AKAP12在HCC中的表达降低,而启动子高甲基化是导致AKAP12表达降低的最重要原因。多种研究结果显示,AKAP12在慢性肝病方面可能是一个潜在的治疗靶点,但尚需进一步研究证实。
此外,AKAP12在慢性乙型肝炎、自身免疫性肝病中的作用尚缺乏研究依据,作用还不明确,需要进一步探讨。
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017